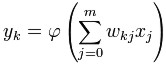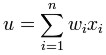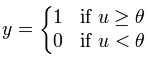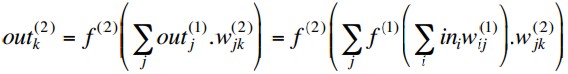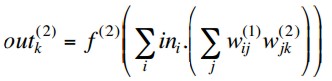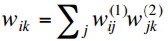Бесплатный учебный инструмент для студентов, преподавателей и
ученых, работающих в областях нейробиологии, биоинформатики, биофизики,
биомедицинской инженерии и искусственного интеллекта.
Краткое содержание
Существующие на сегодняшний день модели искусственных нейронов не
способны моделировать фундаментально важные свойства биологических
нейронов:
1) антагонистические рецептивные поля и
2) выходной ответный сигнал нейрона PSTH на любой стимул (раздражитель).
Даже если некоторые модели нейронов и пытаются моделировать
антагонистические рецептивные поля, то они не способны моделировать
выходной сигнал PSTH, и наоборот – при попытке определенных моделей
моделировать выходной сигнал нейрона PSTH, они не в состоянии
разъяснить происхождение антагонистических рецептивных полей нейронов.
Например, одна из самых популярных моделей DOG (Difference Of
Gaussians) моделирует антагонистическую структуру рецептивного поля,
однако не способна моделировать выходной сигнал нейрона PSTH.
Подавляющее большинство моделей искусственных нейронов не способны
моделировать ни антагонистические рецептивные поля нейронов, ни
выходные сигналы PSTH нейрона.
Впервые в истории нейробиологии модель нейрона RF-PSTH способна
моделировать как антагонистические рецептивные поля нейронов, так и
выходной сигнал PSTH.
Нейромодель RF-PSTH основана на физических свойствах биологических
нейронов.
Рекомендуемая литература
Подробное описание рецептивного поля биологического нейрона изложено в
книге «Encyclopedia
of the Human Brain»,
автор книги: Вилейанур Рамачандран (англ. Vilayanur S. Ramachandran)
(редактор), издательство: «Academic Press», 10 июля 2002 г. (1-ое
издание), ISBN-10: 0122272102, ISBN-13: 978-0122272103.
В этой книге, пожалуйста, прочитайте главу «Receptive Field»,
написанную Rajesh P.
N. Rao (University of Washington), страницы 155-168.
Нейромодель RF-PSTH симулирует функции нейрона, описанные в этой главе.
Значение и преимущества новой нейромодели RF-PSTH
Генерируемый нейромоделью RF-PSTH выходной сигнал PSTH соответствует
данным экспериментальных, лабораторных измерений реальных биологических
нейронов.
Измерения реальных биологических нейронов показывают, что рецептивные
поля сенсорных нейронов имеют симметричную, антагонистическую структуру
концентрических кругов, однако современная наука не может
удовлетворительно объяснить причину этого феномена. Высказываются
гипотеза, что антагонистические, концентрические рецептивные поля
формируются из-за того, что нейрон соединяется с рецепторами (или
другими нейронами) посредством синаптических соединений, и
предположительно эти синаптические соединения распределяются таким
образом, что формируются антагонистические концентрические круги. Не
существует удовлетворительного объяснения того, почему рецептивные поля
должны формировать антагонистические структуры в форме концентрических
кругов.
Нейромодель RF-PSTH может симулировать антагонистические,
концентрические круговые структуры рецептивных полей сенсорных
нейронов.
Нейромодель RF-PSTH утверждает, что антагонистическая круговая
структура рецептивного поля является внутренней особенностью
всех сенсорных нейронов, и для формирования такой структуры не
требуется
никаких внешних нейронных связей (внешних нейронных сетей).
Программа нейромодели
RF-PSTH
По ниже приведенным ссылкам можно скачать программу «Нейромодель
RF-PSTH» v.2.5 для следующих платформ:
Пробную версию Нейромодели RF-PSTH можно свободно использовать
в академических и образовательных целях.
Описание Нейромодели RF-PSTH
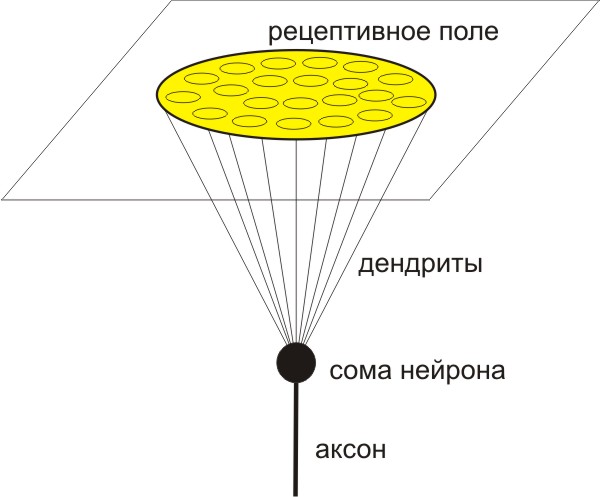
Нейрон имеет тело (сому), дендриты и аксоны.
Нейрон моделируется как трехмерный объект в трехмерном пространстве:
1) сома нейрона моделируется как математическая точка;
2) рецептивное поле нейрона моделируется как двумерная плоскость;
3) дендриты, исходящие из сомы нейрона, достигают двумерной плоскости;
4) в двумерной плоскости дендриты формируют круговое рецептивное поле;
5) трехмерная форма нейрона является прямым круговым конусом, вершина
которого репрезентирует сому нейрона, а основание – рецептивное поле.
Такой вариант хорошо иллюстрируется на примере ганглиозной (нейронной)
клетки, получающей входные сигналы от фоторецепторов в сетчатке.
Большинство других нейронов также довольно неплохо соответствуют этой
модели, например, соматосенсорные нейроны, получающие входные сигналы
от кожи, и т.д.
Рисунок 1. Нейрон моделируется как трехмерный объект в трехмерном
пространстве.
Нейромодель RF-PSTH утверждает, что если выполняются все вышеупомянутые
условия, то тогда нейрон будет иметь симметричную, антагонистическую,
концентрически круговую структуру рецептивного поля. Но если
рецептивное поле нейрона будет иметь другую конфигурацию (к примеру,
будет частью трехмерной сферы, или будет иметь какую-либо другую
конфигурацию), то тогда рецептивное поле нейрона может не иметь
антагонистическую, концентрически круговую структуру. Иными словами,
трехмерная пространственная конфигурация нейрона играет главную роль в
формировании структуры рецептивного поля.
Часто задаваемые вопросы о нейромодели RF-PSTH
Рецептивные поля нейронов могут иметь более сложную структуру,
чем антагонистическая, концентрически круговая структура. Например,
рецептивными полями нейронов V1 являются линии, отрезки или квадратные
формы под некоторым углом и т.д., а нейромодель RF-PSTH не моделирует
такие сложные рецептивные поля нейронов, поэтому я считаю, что
нейромодель RF-PSTH является некорректной/неполной моделью, не так ли?
Нам регулярно раз за разом задают этот вопрос люди, имеющие высокие
академические степени в нейронауке, поэтому ответ таков. Прежде всего,
«рецептивное поле нейрона V1» (так, как оно изображается в статьях и
учебниках по нейронауке) имеет неверное название – фактически оно
является не рецептивным полем одного какого-либо нейрона, а рецептивным
полем многослойной сети (распространяющейся от сетчатки через
латеральное коленчатое ядро (LGN) в область V1). Но многослойная сеть и
один нейрон это две разные вещи. Нейромодель RF-PSTH моделирует
поведение одного нейрона, а не всей многослойной нейросети. Если вы
возьмете действительно только один нейрон зоны V1 (отбросив все
близлежащие нейроны), и если вы измерите входные и выходные
характеристики одного такого нейрона, то получите те же результаты, что
может генерировать и нейромодель RF-PSTH. Другими словами,
недопонимание происходит из-за смешения рецептивного поля одного
нейрона с рецептивным полем многослойной нейросети.
Какие типы нейронов какой части мозга (коры мозга, таламуса,
гипоталамуса, миндалевидного тела, гиппокампа, и т.д.) симулирует
нейромодель RF-PSTH?
Нейромодель RF-PSTH симулирует нейроны, имеющие трехмерную форму
прямого кругового конуса (как показано на рисунке 1). Не имеет значения
в какой части мозга находится такой нейрон, но если он будет иметь
пространственную трехмерную форму прямого кругового конуса, то
нейромодель RF-PSTH сможет симулировать поведение такого нейрона. На
практике нейроны, имеющие форму прямого кругового конуса, легче всего
найти в первом слое входов множественных сенсорных модальностей. Это
хорошо иллюстрируется на примере ганглиозной (нервной) клетки, которая
получает входящие сигналы от фоторецепторов сетчатки. Множество других
нейронов также подходит под этот вариант, например, соматосенсорные
нейроны, которые получают входящие сигналы от рецепторов кожи, и т.д.
Нейромодель RF-PSTH поддерживает гипотезу Вернона Бенджамина Маунткасла
(почетного профессора нейробиологии в Университете Джона Хопкинса),
который заметил, что различные области неокортекса (визуальные,
слуховые и т. д.) очень однородны по внешнему виду и структуре, и
предложил идею о том, что различные области неокортекса одинаково
обрабатывают информацию, выполняя ту же самую основную операцию.
Вернон Бенджамин Маунткасл
(англ. Vernon Benjamin Mountcastle; 15 июля 1918, Шелбивилл, округ
Шелби, Кентукки — 11 января 2015) — невролог из университета Джонса
Хопкинса. Он открыл и характеризовал модульную организацию коры
головного мозга в 1950-х годах. Это открытие стало поворотной точкой в
исследовании коры, настолько, что почти все теории о сенсорных
функциях, возникшие после публикации Маунткасла о соматосенсорной коре,
используют модульную организацию как основу.
Wikipedia
Глава: Организующий принцип
функции мозга – элементарный модуль и распределенная система
В. Маунткасл (Vernon B. Mountcastle)
Выдержка из страниц 56-57:
<....>
Функциональные свойства распределенных систем
Из классической анатомии хорошо известно, что многие крупные
образования головного мозга объединены внешними путями в сложные
системы, включая массивные сети с повторными входами. Три группы
описанных выше недавно обнаруженных фактов пролили новый свет на
системную организацию головного мозга. Первая из них состоит в том, что
основные структуры головного мозга построены по принципу повторения
одинаковых многоклеточных единиц. Эти модули представляют собой
локальные нейронные цепи из сотен или тысяч клеток, объединенных
сложной сетью интрамодульных связей. Модули любого образования более
или менее одинаковы, но в разных образованиях они могут резко
различаться. Модульная единица новой коры — это описанная выше
вертикально организованная группа клеток. Такие основные единицы
представляют собой одиночные трансламинарные цепи нейронов,
мини-колонки, которые в некоторых областях собраны в более крупные
единицы, величина и форма которых неодинаковы в разных местах. Но предполагается, что по своему качественному характеру функция обработки информации в новой коре одинакова в разных областях, хотя этот внутренний аппарат может быть изменен его предыдущей историей, особенно в критические периоды онтогенеза.
Разумный мозг
Эделмен Дж., Маунткасл В.; Алексеенко Н.Ю. (Пер.); Е.Н. Соколов (ред.)
Перевод на русский язык, «Мир», 1981. MIT Press, 1978.
Пошаговые инструкции по запуску программы нейромодели
RF-PSTH
Этап #1
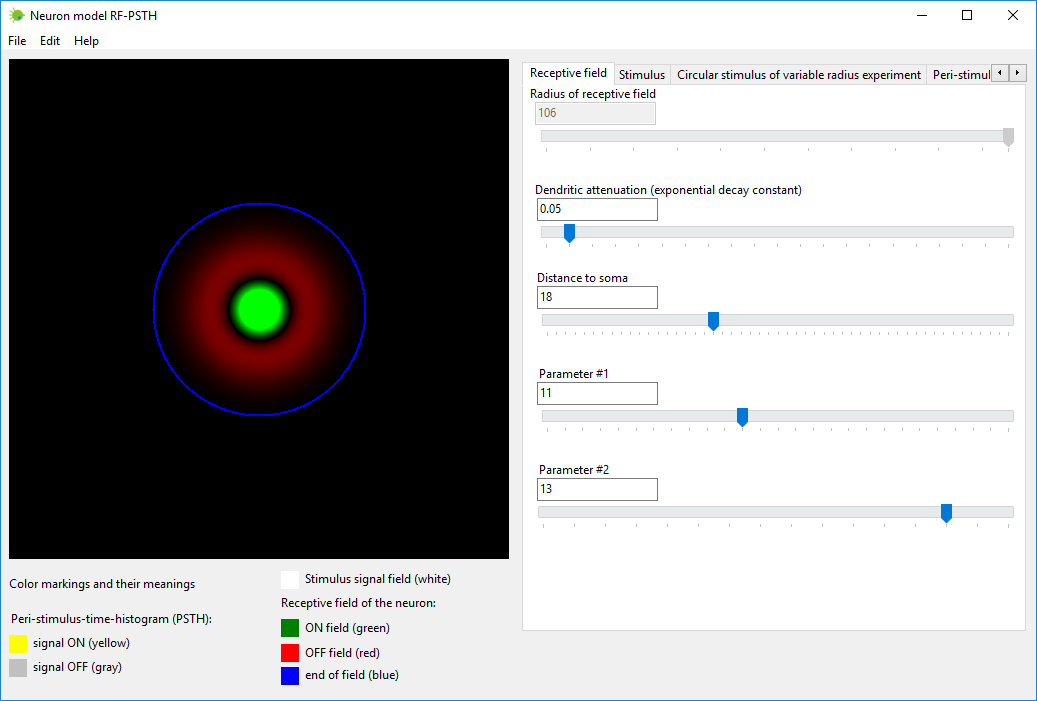
Выберите параметры, определяющие структуру рецептивного поля нейрона.
Их можно выбрать во вкладке под названием «Рецептивное поле»
(“Receptive field”).
Параметры нейрона являются такими: Радиус рецептивного поля (Radius of receptive field) – этот
параметр рассчитывается автоматически,
вам не надо выбирать его значение; Затухание дендритов (константа экспоненциального убывания)
(Dendritic attenuation (exponential decay constant)) – когда
сигнал распространяется по дендритам, он затухает по закону
экспоненциального убывания, при помощи этого параметра устанавливается
константа экспоненциального убывания; Расстояние до сомы (Distance to soma) – расстояние от сомы до
плоскости рецептивного поля; Параметр #1 (Parameter #1) – параметр, устанавливающий диаметр
дендритов; Параметр #2 (Parameter #2) – еще один параметр нейрона.
Рисунок 2. Вкладка «Рецептивное поле» (“Receptive field”) в программе
нейромодели RF-PSTH.
The receptive field of a sensory
neuron is a region of space in which the presence of a
stimulus will alter the firing of that neuron. Receptive fields have
been identified for neurons of the auditory system, the somatosensory
system, and the visual system.
The concept of receptive fields can be extended to further up the
neural system; if many sensory receptors all form synapses with a
single cell further up,
they collectively form the receptive field of that cell. For example,
the
receptive field of a ganglion cell in the retina of the eye is composed
of input from all of the photoreceptors which synapse with it, and a
group of
ganglion cells in turn forms the receptive field for a cell in the
brain. This
process is called convergence.
<...>
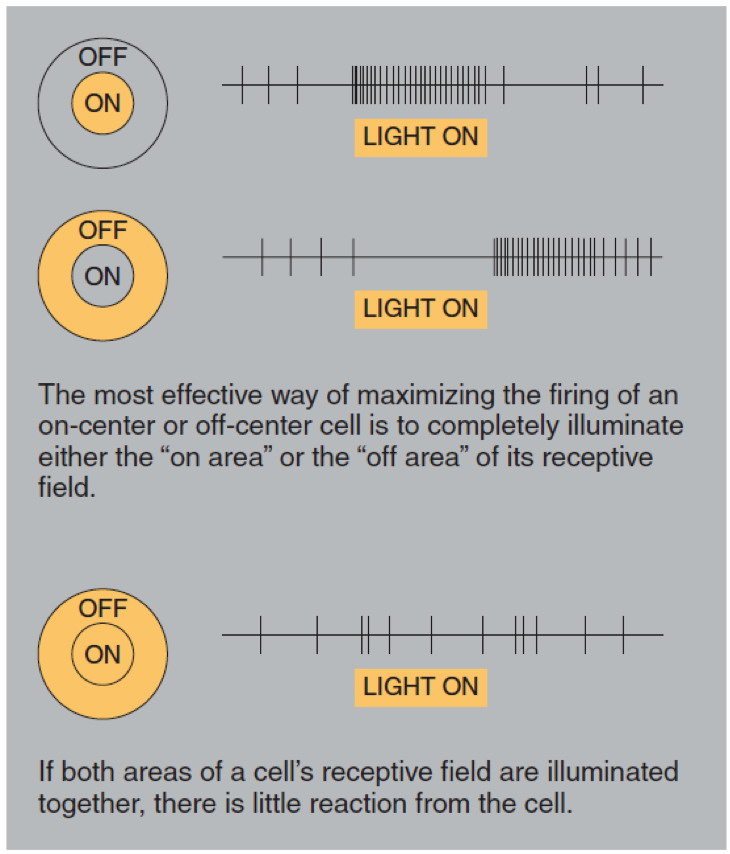
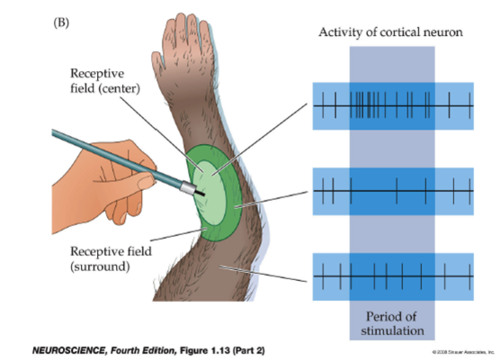
On center and off center retinal ganglion cells respond oppositely to
light in the center and surround of their receptive fields. A strong
response means high frequency firing, a weak response is firing at a
low frequency, and no
response means no action potential is fired.
Wikipedia
Этап #2
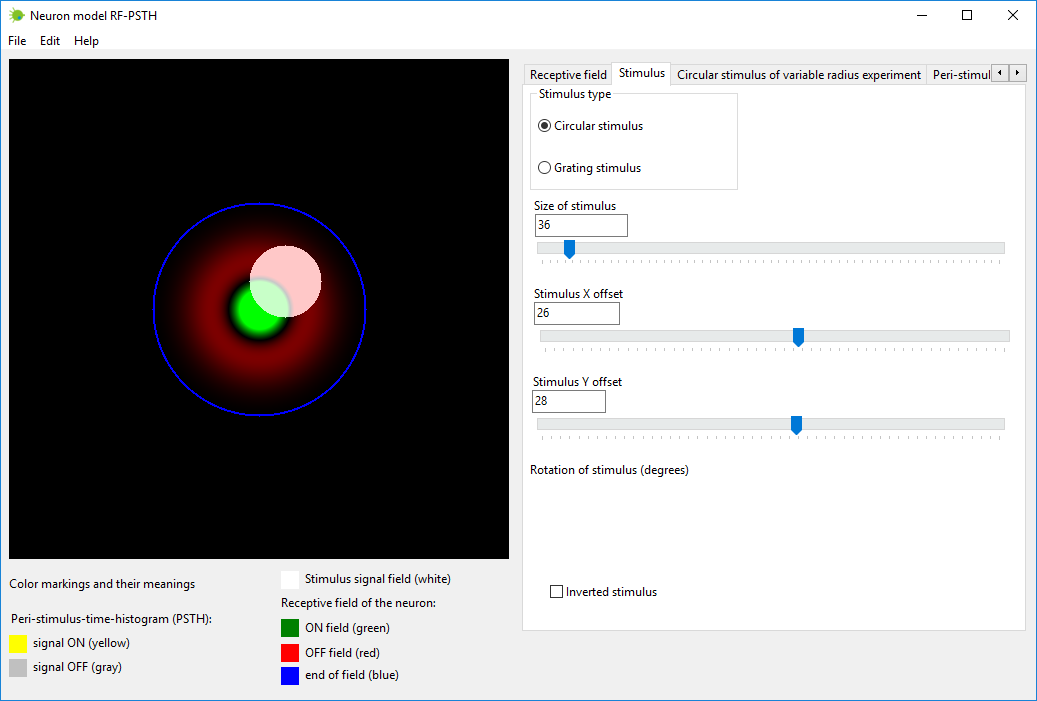
Создайте стимул с нужными параметрами. Это можно осуществить во вкладке
под названием «Стимул» (“Stimulus”).
Стимул изображается белым цветом на рецептивном поле нейрона.
Стимул может быть нескольких типов:
1) круговой стимул (circular stimulus) – вы можете выбрать
радиус и координаты центра круга (x,y);
2) стимул-решетка (grating stimulus) – вы можете выбрать ширину
решетки, координаты параллельного переноса (x,y) и угол поворота (в
градусах);
3) инвертированный стимул (inverted stimulus) – инверсия
стимулированных и не стимулированных областей.
Параметры кругового стимулы можно менять непосредственно мышкой (менять
размер и перемещать), нажав кнопкой мышки на образ стимула.
Стимул-решетку можно изменять только при помощи скользящей метки
(sliding trackbar) в нижеприведенных диапазонах значений.
С помощью нейромодели RF-PSTH также можно симулировать реакцию нейрона
на движущийся стимул, однако в этой версии программы можно симулировать
только статические, не движущиеся стимулы.
Рисунок 3. Вкладка «Стимул» (“Stimulus”) в программе нейромодели
RF-PSTH.
In physiology, a stimulus
(plural stimuli) is a detectable change in the internal or external
environment. The ability of an organism or organ to respond to external
stimuli is called sensitivity.
When a stimulus is applied to a sensory receptor, it normally elicits
or
influences a reflex via stimulus transduction. These sensory receptors
can receive
information from outside the body, as in touch receptors found in the
skin or light
receptors in the eye, as well as from inside the body, as in
chemoreceptors and mechanorceptors.
Wikipedia
Этап #3
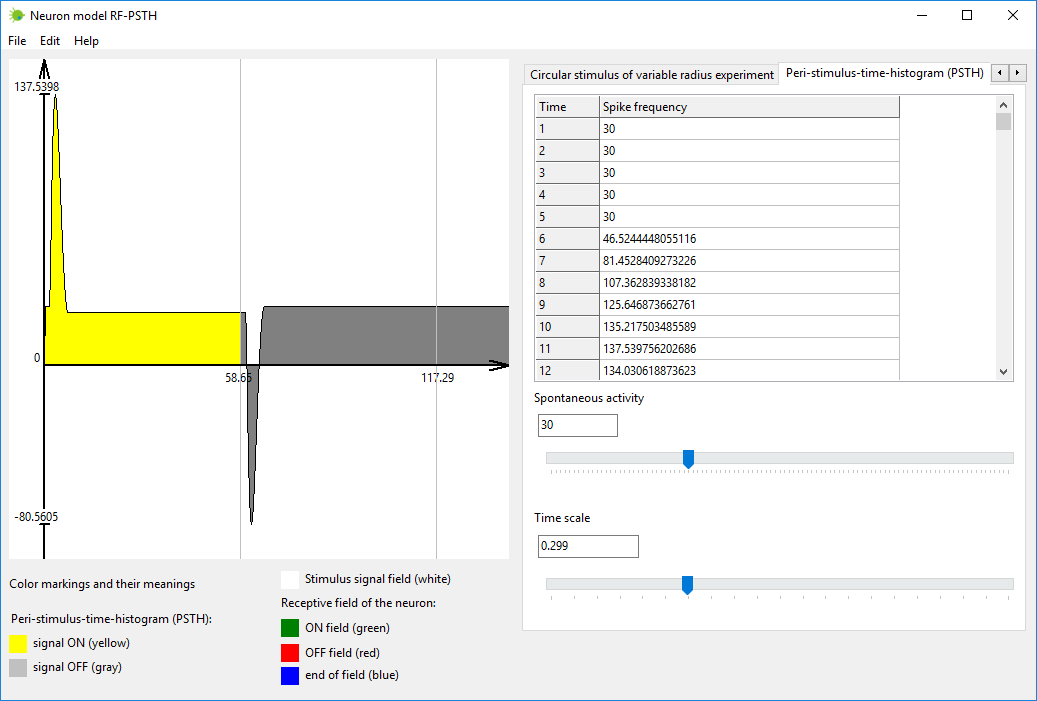
Симулируйте выходной сигнал нейрона как реакцию на входной стимул. Это
можно выполнить во вкладке под названием «Временная перистимульная
гистограмма» (“Peri-stimulus-time-histogram (PSTH)”).
Стимул включается, когда время равно нулю. Стимул выключается
автоматически, когда выходной сигнал нейрона стабилизируется и
становится почти стационарным. Выходной сигнал нейрона отображается
желтым цветом, когда стимул включен, и серым при выключенном стимуле.
Рисунок 4. Вкладка «Временная перистимульная гистограмма»
(“Peri-stimulus-time-histogram (PSTH)”) в программе нейромодели
RF-PSTH.
In neurophysiology, peristimulus
time histogram and poststimulus time histogram, both abbreviated PSTH
or PST histogram, are histograms of the times at which neurons fire.
These histograms are used to visualize
the rate and timing of neuronal spike discharges in relation to an
external stimulus
or event. The peristimulus time histogram is sometimes called perievent
time
histogram, and post-stimulus and peri-stimulus are often hyphenated.
The prefix peri, for through, is typically used in the case of periodic
stimuli, in which case the PSTH show neuron firing times wrapped to one
cycle of the stimulus.
The prefix post is used when the PSTH shows the timing of neuron
firings in response to a stimulus event or onset.
To make a PSTH, a spike train recorded from a single neuron is aligned
with the onset, or a fixed phase point, of an identical stimulus
repeatedly presented to an animal. The aligned sequences are
superimposed in time,
and then used to construct a histogram.
Wikipedia
Дополнительные замечания о выходном сигнале PSTH
Реальные биологические нейроны не могут генерировать потенциалов
действия с отрицательной частотой в своих выходных сигналах. Однако
этот отрицательный выходной сигнал можно измерять как уменьшенный
пресинаптический потенциал внутри сомы нейрона, находящийся в месте,
где аксон соединяется с сомой. Данная симуляционная программа
показывает и раскрывает происходящие в нейроне внутренние процессы.
Выходной сигнал PSTH рассчитывается без учета кратковременной депрессии
(STD) и кратковременной фасилитации (содействия) (STF).
Short-term
plasticity (STP), also called dynamical synapses, refers to a
phenomenon in which synaptic efficacy changes over time in a way that
reflects the history of presynaptic activity. Two types of STP, with
opposite effects on synaptic efficacy, have been observed in
experiments. They are known as short-term depression (STD) and
short-term facilitation (STF). STD is caused by depletion of
neurotransmitters consumed during the synaptic signaling process at the
axon terminal of a pre-synaptic neuron, whereas STF is caused by influx
of calcium into the axon terminal after spike generation, which
increases the release probability of neurotransmitters. STP has been
found in various cortical regions and exhibits great diversity in
properties. Synapses in different cortical areas can have varied forms
of plasticity, being either STD-dominated, STF-dominated, or showing a
mixture of both forms.
Compared with long-term plasticity, which is hypothesized as the neural
substrate for experience-dependent modification of neural circuit, STP
has a shorter time scale, typically on the order of hundreds to
thousands of milliseconds. The modification it induces to synaptic
efficacy is temporary. Without continued presynaptic activity, the
synaptic efficacy will quickly return to its baseline level.
Scholarpedia
Этап #4
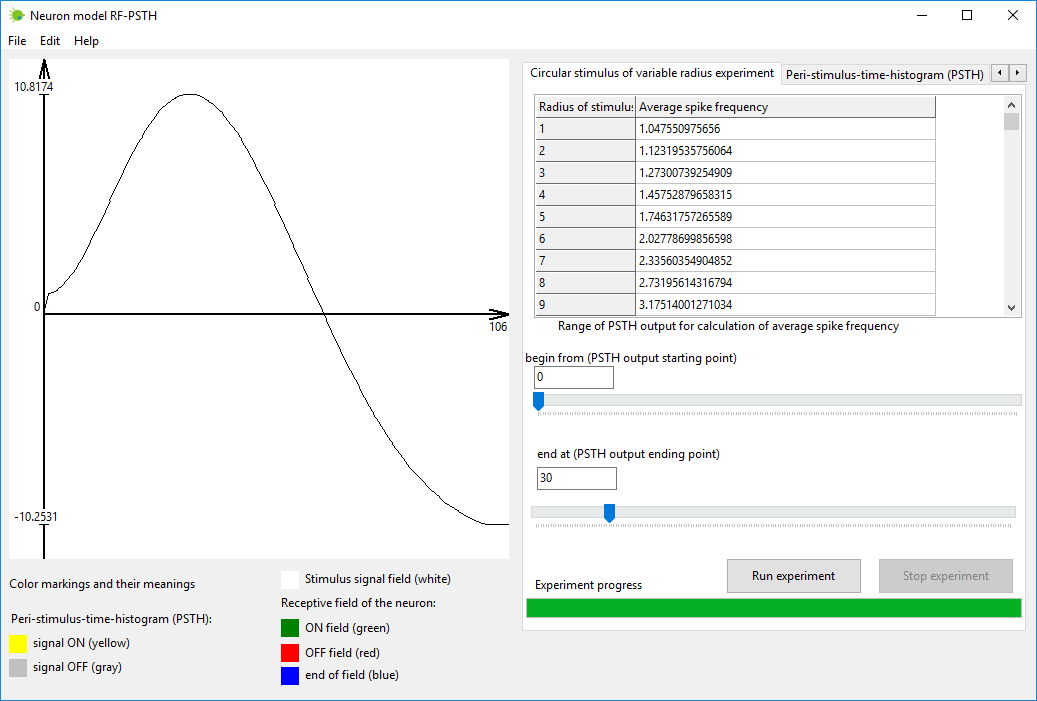
Симулируйте эксперимент с круговым стимулом переменного радиуса. Это
можно выполнить во вкладке «Эксперимент с круговым стимулом переменного
радиуса» (“Circular stimulus of variable radius
experiment”).
Описание эксперимента является следующим. Круговой стимул помещается в
центр рецептивного поля. Размер стимула увеличивается пошагово, начиная
с нуля и заканчивая размером рецептивного поля. На каждом шаге
вычисляется выходной сигнал PSTH. На каждом шаге берется диапазон
выходных значений из графика PSTH и вычисляется средняя частота
потенциалов действия для этого определенного диапазона. Конечный
график, получающийся в результате эксперимента, изображает зависимость
средней частоты потенциалов действия от размера стимула. Форма
конечного графика чувствительна к выбранному диапазону выходного
сигнала PSTH, на основе которого вычисляется средняя частота
потенциалов действия. Изменение выбранного диапазона (выходного сигнала
PSTH) меняет конечный график эксперимента. В нейронауке не существует
правил, которые определяли бы, какой диапазон сигнала PSTH следует
выбрать для вычисления средней частоты потенциалов действия. Поэтому
вам следует самим поэкспериментировать со значениями этого диапазона
для того, чтобы найти такой диапазон, который лучше всего
соответствовал бы вашим целям.
Рисунок 5. Вкладка «Эксперимент с круговым стимулом переменного
радиуса» (“Circular stimulus of variable radius experiment”) в
программе нейромодели RF-PSTH.
Недостатки современных моделей искусственных нейронов и
превосходство нейромодели RF-PSTH
Математическое моделирование любого физического феномена требует
упрощений (редукций) этого феномена для того, чтобы уменьшить
количество моделируемых параметров и количество вычислений. Технический
вопрос состоит в том, насколько данный феномен можно упростить, чтобы
не потерять важную информацию, нужную для решения конкретной задачи.
Например, если нам нужно проанализировать движение автомобиля по
дороге, мы можем редуцировать автомобиль до математической точки (не
имеющей размеров и массы), которая движется через двумерное
пространство с некоторой скоростью. Такая упрощенная модель автомобиля
годится для того, если мы хотим, к примеру, рассчитать, сколько времени
у автомобиля уйдет на переезд из точки А в точку Б. Однако, если нам
надо выяснить, какая сила воздействует на тормозные диски автомобиля,
когда водитель нажимает на тормозные педали для остановки машины, то
тогда модели, в которой автомобиль отображается как математическая
точка, будет недостаточно для решения такой задачи. Если вам требуется
выяснить, какая сила действует на тормозные диски автомобиля, когда
водитель нажимает на тормозную педаль, то тогда вам надо знать массу
автомобиля, диаметр колес, и т.д. – но все эти параметры были исключены
из описанной выше модели автомобиля. Другими словами, при создании
математической модели можно исключить некоторые существенные
особенности (параметры), без которых будет невозможно решить некоторые
специфические практические задачи.
Рассмотрим более подробно используемые на сегодняшний день модели
искусственных нейронов.
The first artificial neuron was the Threshold Logic Unit (TLU) first
proposed by Warren McCulloch and Walter Pitts in 1943. As a transfer
function, it employed a threshold, equivalent to using the Heaviside
step function. Initially, only a simple model was considered, with
binary inputs and outputs, some restrictions on the possible weights,
and a more flexible threshold value. Since the beginning it was already
noticed that any boolean function could be implemented by networks of
such devices, what is easily seen from the fact that one can implement
the AND and OR functions, and use them in the disjunctive or the
conjunctive normal form.
Researchers also soon realized that cyclic networks, with feedbacks
through neurons, could define dynamical systems with memory, but most
of the research concentrated (and still does) on strictly feed-forward
networks because of the smaller difficulty they present.
One important and pioneering artificial neural network that used the
linear threshold function was the perceptron, developed by Frank
Rosenblatt. This model already considered more flexible weight values
in the neurons, and was used in machines with adaptive capabilities.
The representation of the threshold values as a bias term was
introduced by Bernard Widrow in 1960.
In the late 1980s, when research on neural networks regained strength,
neurons with more continuous shapes started to be considered. The
possibility of differentiating the activation function allows the
direct use of the gradient descent and other optimization algorithms
for the adjustment of the weights. Neural networks also started to be
used as a general function approximation model.
<...> Basic structure
For a given artificial neuron, let there be m + 1 inputs with signals x0
through xm and weights w0 through wm.
Usually, the x0 input is assigned the value +1,
which makes it a bias input with wk0 = bk.
This leaves only m actual
inputs to the neuron: from x1 to xm.
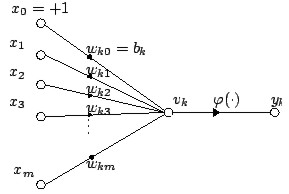
The output of kth neuron is:
Where φ is the transfer
function.
The output is analogous to the axon of a biological neuron, and its
value propagates to input of the next layer, through a synapse. It may
also exit the system, possibly as part of an output vector.
It has no learning process as such. Its transfer function weights are
calculated and threshold value are predetermined.
<...> Types of transfer functions
The transfer function of a neuron is chosen to have a number of
properties which either enhance or simplify the network containing the
neuron. Crucially, for instance, any multilayer perceptron using a
linear transfer function has an equivalent single-layer network; a
non-linear function is therefore necessary to gain the advantages of a
multi-layer network.
Below, u refers in all cases to the weighted sum of all the inputs to
the neuron, i.e. for n inputs,
where w is a vector of
synaptic weights and x is a
vector of inputs.
Step function
The output y of this transfer function is binary, depending on whether
the input meets a specified threshold, θ. The "signal" is sent, i.e. the
output is set to one, if the activation meets the threshold.
This function is used in perceptrons and often shows up in many other
models. It performs a division of the space of inputs by a hyperplane.
It is specially useful in the last layer of a network intended to
perform binary classification of the inputs. It can be approximated
from other sigmoidal functions by assigning large values to the weights.
Linear combination
In this case, the output unit is simply the weighted sum of its inputs
plus a bias term. A number of such linear neurons perform a linear
transformation of the input vector. This is usually more useful in the
first layers of a network. A number of analysis tools exist based on
linear models, such as harmonic analysis, and they can all be used in
neural networks with this linear neuron. The bias term allows us to
make affine transformations to the data.
Sigmoid
A fairly simple non-linear function, a Sigmoid function such as the
logistic function also has an easily calculated derivative, which can
be important when calculating the weight updates in the network. It
thus makes the network more easily manipulable mathematically, and was
attractive to early computer scientists who needed to minimize the
computational load of their simulations. It is commonly seen in
multilayer perceptrons using a backpropagation algorithm.
<...> Comparison to biological neurons
Artificial neurons bear a striking similarity to their biological
counterparts.
Dendrites - In a biological neuron, the dendrites act as the input
vector. These dendrites allow the cell to receive signals from a large
(>1000) number of neighboring neurons. As in the above mathematical
treatment, each dendrite is able to perform "multiplication" by that
dendrite's "weight value." The multiplication is accomplished by
increasing or decreasing the ratio of synaptic neurotransmitters to
signal chemicals introduced into the dendrite in response to the
synaptic neurotransmitter. A negative multiplication effect can be
achieved by transmitting signal inhibitors (i.e. oppositely charged
ions) along the dendrite in response to the reception of synaptic
neurotransmitters.
Soma - In a biological neuron, the soma acts as the summation function,
seen in the above mathematical description. As positive and negative
signals (exciting and inhibiting, respectively) arrive in the soma from
the dendrites, the positive and negative ions are effectively added in
summation, by simple virtue of being mixed together in the solution
inside the cell's body.
Axon - The axon gets its signal from the summation behavior which
occurs inside the soma. The opening to the axon essentially samples the
electrical potential of the solution inside the soma. Once the soma
reaches a certain potential, the axon will transmit an all-in signal
pulse down its length. In this regard, the axon behaves as the ability
for us to connect our artificial neuron to other artificial neurons.
Unlike most artificial neurons, however, biological neurons fire in
discrete pulses. Each time the electrical potential inside the soma
reaches a certain threshold, a pulse is transmitted down the axon. This
pulsing can be translated into continuous values. The rate (activations
per second, etc.) at which an axon fires converts directly into the
rate at which neighboring cells get signal ions introduced into them.
The faster a biological neuron fires, the faster nearby neurons
accumulate electrical potential (or lose electrical potential,
depending on the "weighting" of the dendrite that connects to the
neuron that fired). It is this conversion that allows computer
scientists and mathematicians to simulate biological neural networks
using artificial neurons which can output distinct values (often from
-1 to 1).
Wikipedia
Все эти модели искусственных нейронов не могут объяснить и
симулировать антагонистические рецептивные поля и выходной сигнал PSTH
реальных
биологических нейронов. На основе таких недействительных искусственных
нейронов строятся Искусственные Нейронные Сети.
Если Искусственная Нейронная Сеть (ИНС) строится из нейронов с линейной
передаточной функцией, то тогда такую многослойную сеть можно упростить
до однослойной сети, которая будет эквивалентна исходной многослойной
сети. Поэтому нелинейная передаточная функция создает преимущества для
многослойной сети.
We have noted before that if we have a regression problem with
non-binary network outputs, then it is appropriate to have a linear
output activation function. So why not simply use linear activation
functions on the hidden layers as well?
With activation functions f(n)(x)
at layer n, the outputs of a
two-layer MLP are
so if the hidden layer activations are linear, i.e. f(1)(x) = x, this simplifies to
But this is equivalent to a single layer network with weights
and we know that such a network cannot deal with non-linearly separable
problems.
Learning in Multi-Layer
Perceptrons - Back-Propagation
Neural Computation : Lecture 7
John A. Bullinaria, 2013
Если в построении искусственного нейрона будет использоваться
линейная передаточная функция, и если количество входных нейронных
сигналов
будет сведено до одного(1) входного сигнала, то тогда способности
обработки информации у такого нейрона упадут почти до нуля. Этот пример
ясно показывает, что чрезмерное упрощение реального физического объекта
сделает математическую модель неспособной решать практические проблемы.
Информатики, которые работают с искусственными нейронными сетями,
предпочитают использовать:
1) нейроны с нелинейными передаточными функциями, при этом полагая, что
нелинейный перцептрон лучше линейного;
2) многослойные сети с нелинейными передаточными функциями, т.к. было
доказано, что однослойную сеть нельзя научить распознаванию многих
классов образов.
The perceptron
algorithm was invented in 1957 at the Cornell Aeronautical Laboratory
by Frank Rosenblatt.
<...>
Although the perceptron initially seemed promising, it was eventually
proved that perceptrons could not be trained to recognize many classes
of patterns. This led to the field of neural network research
stagnating for many years, before it was recognised that a feedforward
neural network with two or more layers (also called a multilayer
perceptron) had far greater processing power than perceptrons with one
layer (also called a single layer perceptron). Single layer perceptrons
are only capable of learning linearly separable patterns; in 1969 a
famous book entitled Perceptrons by Marvin Minsky and Seymour Papert
showed that it was impossible for these classes of network to learn an
XOR function. It is often believed that they also conjectured
(incorrectly) that a similar result would hold for a multi-layer
perceptron network. However, this is not true, as both Minsky and
Papert already knew that multi-layer perceptrons were capable of
producing an XOR Function. <...> Three years later Stephen
Grossberg published a series of papers introducing networks capable of
modelling differential, contrast-enhancing and XOR functions. (The
papers were published in 1972 and 1973, see e.g.: Grossberg, Contour
enhancement, short-term memory, and constancies in reverberating neural
networks. Studies in Applied Mathematics, 52 (1973), 213-257, online).
Nevertheless the often-miscited Minsky/Papert text caused a
significant decline in interest and funding of neural network research.
It took ten more years until neural network research experienced a
resurgence in the 1980s. This text was reprinted in 1987 as
"Perceptrons - Expanded Edition" where some errors in the original text
are shown and corrected.
The kernel perceptron algorithm was already introduced in 1964 by
Aizerman et al. Margin bounds guarantees were given for the
Perceptron algorithm in the general non-separable case first by Freund
and Schapire (1998), and more recently by Mohri and Rostamizadeh
(2013) who extend previous results and give new L1 bounds.
Wikipedia
Информатики не способны заметить одну существенную фундаментальную
особенность перцептрона – когда перцептрон используется для задач
распознавания образов, то перцептрон действует как простая техника
соответствия шаблонов (template matching), и не имеет значения,
является ли передаточная функция перцептрона линейной или нелинейной.
Нелинейность в этом случае совсем ничего не дает – перцептрон ведет
себя как простая техника соответствия шаблонов. Общеизвестным фактом
является то, что техника соответствия шаблонов не способна распознать
объект при изменении его размеров, повороте или перемещении. Иначе
говоря, информатики не способны понять очевидный факт того, что
перцептрон фундаментально неспособен распознать объект, который меняет
форму (меняет размер, вращается или перемещается).
Когда нелинейный перцептрон учат фильтровать аудио сигналы, и для
обучения такого перцептрона используется техника обучения по методу
обратного распространения ошибки, то результаты экспериментов
показывают, что, когда количество итераций обучения стремится к
бесконечности, то веса перцептрона стремятся к коеффициентам DSP
фильтра, который является оптимальным для фильтрации данного
конкретного сигнала. Иначе говоря, не имеет значения, является ли
передаточная функция у перцептрона линейной или нелинейной, –
перцептрон действует как простой линейный DSP фильтр (цифровой
обработчик сигналов). Нелинейность передаточной функции перцептрона не
придает ему никаких преимуществ перед линейным DSP фильтром.
Информатики, работающие с искусственными нейронными сетями, используют
слишком упрощенные математические модели нейрона, которые не способны
решать практические проблемы по распознаванию образов. Когда был
изобретен искусственный нейрон, то делались утверждения, что вскоре у
человечества будут машины, способные решать те же задачи по
распознаванию образов, которые может решать и биологический мозг.
Однако, несмотря на десятилетия исследований и нудной работы,
нелинейные многослойные перцептроны все еще неспособны решать даже
самые примитивные задачи по распознаванию образов. Причина такой
неудачи очевидна – модели искусственных нейронов являются слишком
упрощенными, в них недостает некоторых существенных особенностей
реальных биологических нейронов. К примеру, в этих нейронных моделях
недостает антагонистической структуры рецептивного поля, и т.д.
Как правило, информатики не имеют никакого понятия о том, что такое
«рецептивное поле», и уровень их знаний об особенностях и параметрах
реальных биологических нейронов почти нулевой – они не знают, что
рецептивные поля сенсорных нейронов имеют симметричную и
антагонистическую структуру концентрических кругов, и т.д.
Нейромодель RF-PSTH возмещает недостатки и ограничения современных
моделей искусственных нейронов.
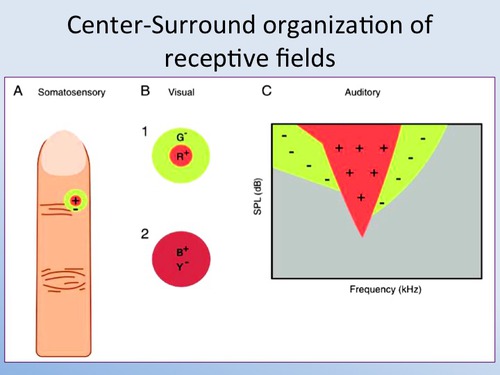
Структура рецептивного поля реальных биологических нейронов
Рецептивные поля сенсорных нейронов имеют симметричную и
антагонистическую структуру концентрических кругов. Ниже приводится
несколько примеров из учебников по нейронауке.
The receptive fields of bipolar
cells are circular. But the centre and the
surrounding area of each circle work in opposite ways: a ray of light
that strikes the centre of the field has the opposite effect from one
that strikes the area surrounding it (known as the "surround").
In fact, there are two types of bipolar cells, distinguished by the way
they respond to light on the centers of their receptive fields. They
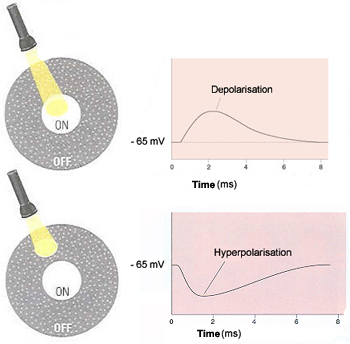
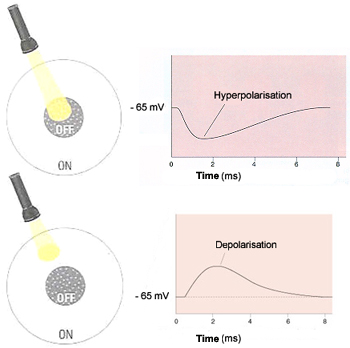
are called ON-centre cells and OFF-centre cells.
If a light stimulus applied to the centre of a bipolar cell's
receptive field has an excitatory effect on that cell, causing it to
become depolarized, it is an ON-centre cell. A ray of light that falls
only on the surround, however, will have the opposite effect on such a
cell, inhibiting (hyperpolarizing) it.
The other kind of bipolar cells, OFF-centre cells,
display exactly the reverse behavior: light on the field's centre has
an inhibitory (hyperpolarizing) effect, while light on the surround has
an excitatory (depolarizing ) effect.
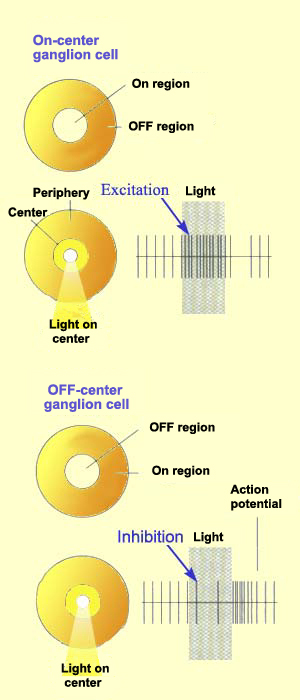
Just like bipolar cells, ganglion cells have concentric receptive
fields with a centre-surround antagonism. But contrary to the two types
of bipolar cells, ON-centre ganglion cells and OFF-centre ganglion
cells do not respond by depolarizing or hyperpolarizing, but rather by
increasing or decreasing the frequency with which they discharge action
potentials.
That said, the response to the stimulation of the centre of the
receptive field is always inhibited by the stimulation of the surround.
Olfactory and Gustatory
Receptive Fields
<...>
olfactory receptive fields are analogous to retinal center-surround
receptive fields: Mitral cells in the olfactory bulb exhibit excitatory
responses to certain chemical compounds in a homologous series of
compounds and inhibitory responses to neighboring compounds that flank
the excitatory compounds in the series. The presence of antagonistic
center-surround receptive fields in the olfactory system suggests that
higher level receptive fields, such as those of neurons in the
olfactory cortex, may possess an oriented structure analogous to visual
and auditory cortical receptive fields. However, the orientation would
be in the space of chemical concentration and time, implying a
sensitivity toward increasing or decreasing amounts of particular
chemical compounds at a particular rate.
Encyclopedia of The Human Brain
- Vol. I, II, III and IV (2002). Editor-in-Chief: V. S. Ramachandran.
ISBN: 978-0-12-227210-3.
Volume IV, Page 165
On-center and
Off-center receptive fields. The receptive fields of retinal ganglion
cells and thalamic neurons are
organized as two concentric circles with different contrast polarities.
On-center neurons respond to the presentation of a light spot on a dark
background and off-center
neurons to the presentation of a dark spot on a light background.
Scholarpedia
Ответная реакция реальных биологических нейронов на стимул
Ниже приводится несколько примеров данных экспериментальных измерений
реальных биологических нейронов.
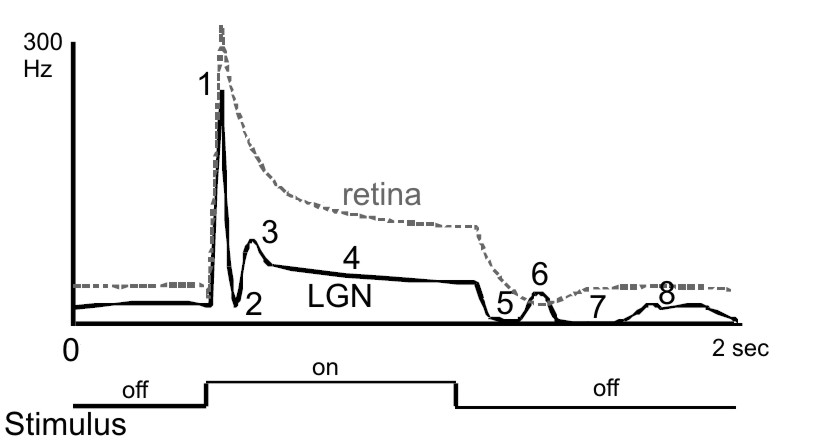
Figure 6. Typical LGN cell responses. The peri-stimulus-time histogram
(PSTH) on top shows a typical temporal waveform of a geniculate (thick
line) and retinal (broken line) visual response to a light spot flashed
on and off within the center of the receptive field. The response to a
sudden increment and decrement of RF illumination can show up to 8
components: 1) initial transient response (overshoot, peak), 2)
post-peak inhibition, 3) early rebound response, 4) tonic response, 5)
stimulus off inhibition (off-response), 6) first post-inhibitory
rebound, 7) late inhibitory response, and 8) second post-inhibitory
rebound. The response profile of the retinal input is less complex.
F Wörgötter, K Suder, N
Pugeault, and K Funke (2003)
Response characteristics in the lateral geniculate nucleus and their
primary afferent influences on the visual cortex of cat
Modulation of Neuronal Responses: Implications for Active Vision. (G T
Buracas, O Ruksenas, G M Boyton and T D Albright, eds.) NATO Science
Series 1: Life and Behavioral Sciences 334:165–188.
Page 170
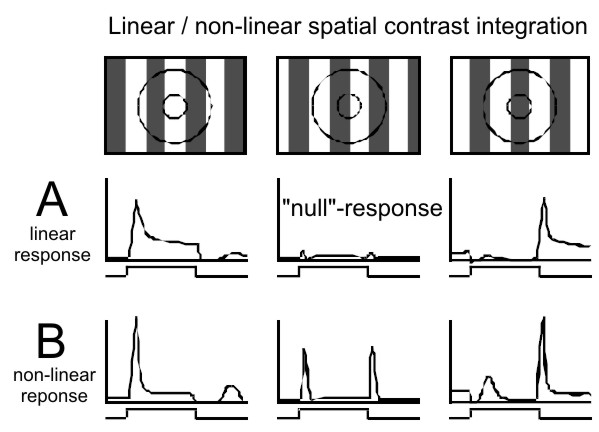
Figure 7. Linear, X-type (A) and nonlinear, Y-type (B) spatial contrast
integration. A) The strongest visual responses of the linear type are
elicited in LGN X-cells by a contrast pattern of a spatial frequency
that fits well to the diameter of the center of the RF. The strength of
the visual response depends on the spatial phase of the pattern (e.g. a
grating). A balanced stimulation of the RF center (and surround) by
bright and dark bars results in the null-response (middle) which is
characterized by only small, if any, change in activity. B) Y-cell
activity is more phasic and is also characterized by the lack of a
null-response. Non-linear (second order) response peaks are observed
irrespective of the spatial frequency of the stimulus.
F Wörgötter, K Suder, N
Pugeault, and K Funke (2003)
Response characteristics in the lateral geniculate nucleus and their
primary afferent influences on the visual cortex of cat
Modulation of Neuronal Responses: Implications for Active Vision. (G T
Buracas, O Ruksenas, G M Boyton and T D Albright, eds.) NATO Science
Series 1: Life and Behavioral Sciences 334:165–188.
Page 172
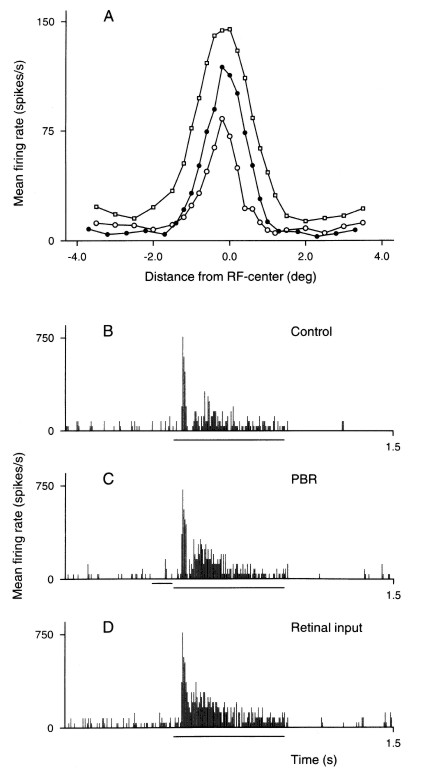
Figure 8. Effect of electrical PBR-stimulation on
the spatial receptive field profile for an on-centre nonlagged Y-cell.
A. Responses to a light slit presented in different position across the
receptive field. The width of the slit was about 1/3 of the width of
the handplotted receptive field centre, the slit length about 3 times
the diameter of the hand-plotted receptive field centre. Each data
point is the average of 5 stimulus presentations. Open circles,
response of the dLGN cell in the control condition, open squares,
retinal input measured by S-potentials. Filled spots, response of dLGN
cell to electrical stimulation of PBR (140 Hz for 70 ms). B-D: Example
of response patterns for the dLGN cell response in the control
condition, with PBR-stimulation, and for the retinal input. The long
horizontal line below the abcissa marks the period when the stimulus
was on, the short bar marks the period with electrical PBR stimulation.
Paul Heggelund (2003)
Signal Processing in the Dorsal Lateral Geniculate Nucleus
Modulation of Neuronal Responses: Implications for Active Vision. (G T
Buracas, O Ruksenas, G M Boyton and T D Albright, eds.) NATO Science
Series 1: Life and Behavioral Sciences 334:109–134.
Page 129
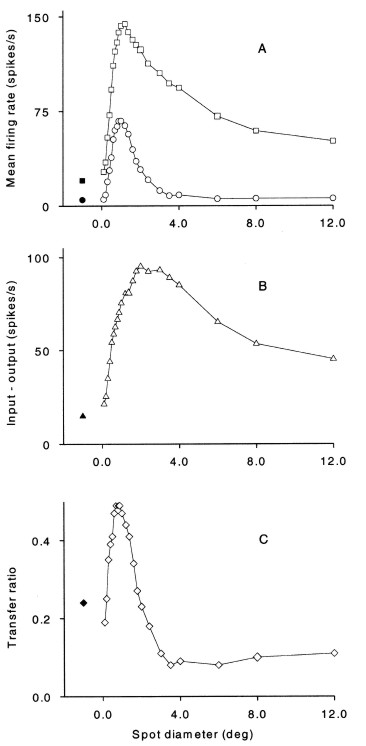
Figure 9. Comparison of the response of an
on-centre X-cell and the response in its retinal input. The stimulus
was a circular light spot of variable diameter but fixed contrast,
centered on the receptive field. A. Open squares, retinal input, open
circles,
dLGN cell response. The response was measured as the average firing
rate during a 500 ms period of spot presentation. Each point is the
average from 20 stimulus presentations. Retinal input was measured as
average frequency of S-potentials. The filled symbols mark spontaneous
activity. B. The difference between retinal input and dLGN cell
response. The data show the reduction in the average response to the
various spot diameters. C. Transfer ratio calculated as output /input
firing rate.
Paul Heggelund (2003)
Signal Processing in the Dorsal Lateral Geniculate Nucleus
Modulation of Neuronal Responses: Implications for Active Vision. (G T
Buracas, O Ruksenas, G M Boyton and T D Albright, eds.) NATO Science
Series 1: Life and Behavioral Sciences 334:109–134.
Page 119
«Нейромодель RF-PSTH» и «Нейрокластерная Модель Мозга» – это две
разные, не связанные между собой модели.
Истинность или ошибочность «Нейромодели RF-PSTH» никак не связанна с
истинностью или ошибочностью «Нейрокластерной Модели Мозга».